Naturaliste du N.-B. en vedette : Nouveaux ténias infectant les poissons du Nouveau-Brunswick et la nécessité d’une science citoyenne !
De Megan Fraser
Les parasites sont des organismes qui vivent sur ou à l’intérieur d’un organisme hôte ; l’hôte subit des dommages et le parasite en profite (Loker & Hofkin 2015). Le parasitisme est un mode de vie symbiotique commun et les parasites sont présents dans tous les règnes de la vie, sous de nombreuses formes et tailles. Presque toutes les plantes et les animaux sont les hôtes d’un ou de plusieurs organismes parasitaires. L’étude des parasites et de leurs hôtes est connue sous le nom de parasitologie, et l’étude des vers parasites en particulier est appelée helminthologie. Les vers parasites appartiennent à un groupe de parasites appelé « helminthes », qui comprend des classes telles que les nématodes (vers ronds), les trématodes (douves) et les cestodes (ténias). Les helminthes peuvent être des ectoparasites, spécialisés dans la vie à l’extérieur de leur hôte, ou des endoparasites, qui vivent à l’intérieur de leur hôte. Pour le non-initié, le mot « ténia » évoque de nombreuses émotions, principalement le dégoût et la répulsion. Aussi étranges et dérangeants que soient les ténias, ils sont pas mal fascinants. Non seulement les ténias privent leur hôte de leurs nutriments, mais certaines espèces modifient également le comportement, le système immunitaire et la capacité de reproduction de leur hôte. Les poissons ne sont pas étrangers aux infections helminthiques, puisqu’on estime que 30 000 espèces d’helminthes infectent les poissons dans le monde entier (Jyrwa et al. 2016) !
J’ai eu l’occasion de me plonger dans le domaine de l’helminthologie après que plusieurs ténias prélevés sur des poissons du Nouveau-Brunswick ont été envoyés au parasitologue Michael Duffy (UNBF) et m’ont été envoyés pour identification. L’identification des espèces de ténias est difficile, surtout au stade larvaire. Les ténias ont peu de caractéristiques morphologiques distinctives, et certaines espèces infectent plusieurs poissons hôtes, de sorte que l’identité de l’hôte ne peut pas être utilisée pour l’identification spécifique du cestode. L’identification au niveau de l’espèce est encore plus compliquée par le fait que de nombreux ténias n’ont pas encore été découverts et nommés !


Des ténias ont récemment été prélevés sur des poissons provenant de deux endroits distincts dans le Nouveau-Brunswick (Fraser et al. 2023). Le premier groupe a été prélevé en 2017 sur des chabots visqueux dans le nord du Nouveau-Brunswick, au cours d’enquêtes non reliées sur les impacts de la gestion forestière sur la santé des poissons (White 2020, Negrazis et al. 2022). Le deuxième ensemble a été recueilli en 2021, après avoir été libéré de son hôte au cours d’une étude sur les poissons dans le sud du Nouveau-Brunswick. D’autres de ces ténias ont été prélevés sur des naseux noirs au même endroit en 2022. En comparant les spécimens de parasites du nord et du sud, deux caractéristiques morphologiques étaient évidentes : les vers du nord du Nouveau-Brunswick étaient segmentés et relativement courts (figure 1), tandis que les vers du sud du Nouveau-Brunswick n’étaient pas segmentés et étaient relativement longs (figure 2). En raison de ces différences morphologiques, je savais que j’avais en main deux espèces distinctes de parasites. J’ai donc prélevé un petit morceau de l’extrémité de chaque ver et j’en ai extrait l’ADN. En comparant l’ADN à celui de tous les ténias connus disponibles dans une grande base de données de séquences génétiques (GenBank), j’ai fait deux découvertes :
1. Les vers du nord du Nouveau-Brunswick ont 89-92% d’identité partagée avec les ténias du genre Schistocephalus, ce qui confirme que ces vers appartiennent au genre Schistocephalus et qu’ils constituent potentiellement une nouvelle espèce.
2. Les vers du sud du Nouveau-Brunswick présentent une identité de plus de 99 % avec le ténia Ligula intestinalis, ce qui a permis l’identification définitive de l’espèce de ce parasite. Ligula intestinalis n’avait pas été enregistré auparavant au Nouveau-Brunswick.
Les espèces de Schistocephalus sont généralement considérées comme des spécialistes de l’épinoche à trois épines et de l’épinoche à neuf épines, mais les vers du nord du Nouveau-Brunswick ont été trouvés dans le chabot visqueux. Il semble qu’un changement d’hôte et un événement de spéciation non documentés se soient produits dans le genre Schistocephalus, donnant naissance aux vers trouvés dans le chabot mince du nord du Nouveau-Brunswick. Il s’agit potentiellement d’une nouvelle espèce et d’une découverte scientifique très intéressante !
L’absence de rapports précédents sur L. intestinalis au Nouveau-Brunswick est notable. Notre identification récente soulève la question de savoir si ce ténia a été récemment introduit ou s’il a été présent pendant de nombreuses années sans être identifié. Ligula intestinalis a trois hôtes au cours de son cycle de vie (Arme & Owen 1968, Szalai et al. 1989, Loot et al. 2002, Biswas & Ash 2021). Elle se reproduit dans l’intestin d’un oiseau piscivore et les œufs passent avec les fèces de l’oiseau dans de l’eau douce. Les œufs éclosent avant d’être mangés par de minuscules crustacés appelés « copépodes » ; les copépodes infectés sont mangés par les poissons, les poissons infectés sont mangés par les oiseaux, et le cycle de vie se répète. Les vers Ligula intestinalis peuvent devenir très grands, atteignant jusqu’à 100 cm de long (Yoneva et al. 2015) ! L’introduction récente de L. intestinalis au Nouveau-Brunswick est possible, surtout en raison de la nature très mobile de l’oiseau hôte. Cependant, il est également possible que L. intestinalis n’ait pas été identifié au Nouveau-Brunswick en raison d’un manque d’intérêt pour la recherche sur les cestodes. Cette question reste sans réponse.
Ligula intestinalis peut « castrer parasitairement » son poisson hôte. En d’autres termes, l’infection limite fortement la capacité des poissons infectés à se reproduire. Cet effet de castration est signalé chez de nombreuses espèces européennes de poissons, notamment le chevaine, le gardon, le rotengle, la brème, le méné et la vandoise, mais il n’a pas été bien étudié dans les populations de poissons d’Amérique du Nord. Les poissons infectés peuvent présenter une croissance gonadique faible ou nulle, des tubercules de ponte moins proéminents, des niveaux d’hormones sexuelles significativement plus faibles, et peuvent présenter un comportement anormal pendant leur période de ponte (Orr 1966, Arme & Owen 1968, Carter et al. 2005, Hecker & Karbe 2005, Schabuss 2005, Trubiroha et al. 2009, Trubiroha et al. 2010). Le mécanisme qui provoque la castration du poisson hôte est inconnu, mais on suppose qu’il s’agit d’une protéine ou d’une autre molécule activement sécrétée par le parasite ! En laboratoire, mes prochaines étapes consisteront à collecter les sécrétions de ces ténias et à identifier les molécules principales sécrétées. L’identification de ces molécules nous rapprochera, je l’espère, d’une meilleure compréhension des mécanismes qui conduisent à la castration parasitaire de leur poisson hôte.
Je cherche à déterminer l’aire de distribution géographique de L. intestinalis en surveillant les populations naturelles de poissons au Nouveau-Brunswick ; ce parasite est-il limité aux lacs du sud du Nouveau-Brunswick ou est-il présent dans l’ensemble de la province ? Il est important de connaître l’aire de distribution géographique donné les conséquences négatives potentielles des infections par L. intestinalis sur la reproduction de l’hôte, en particulier dans les populations de poissons menacées. C’est là que j’ai décidé qu’une initiative de science citoyenne serait utile ! Les pêcheurs de loisir trouvent souvent des ténias dans les poissons sauvages. En faisant participer le public, j’espère que les pêcheurs récréatifs conserveront les ténias qu’ils trouvent afin que je puisse les collecter pour les identifier. Si des spécimens correspondent à L. intestinalis, cela nous donnera des informations précieuses sur l’aire de distribution du parasite au Nouveau-Brunswick et sur les espèces de poissons qui peuvent servir d’hôtes. Veuillez contacter Megan Fraser (courriel : mfrase12@unb.ca) si vous trouvez des ténias pendant que vous pêchez, et la collecte des spécimens sera organisée !
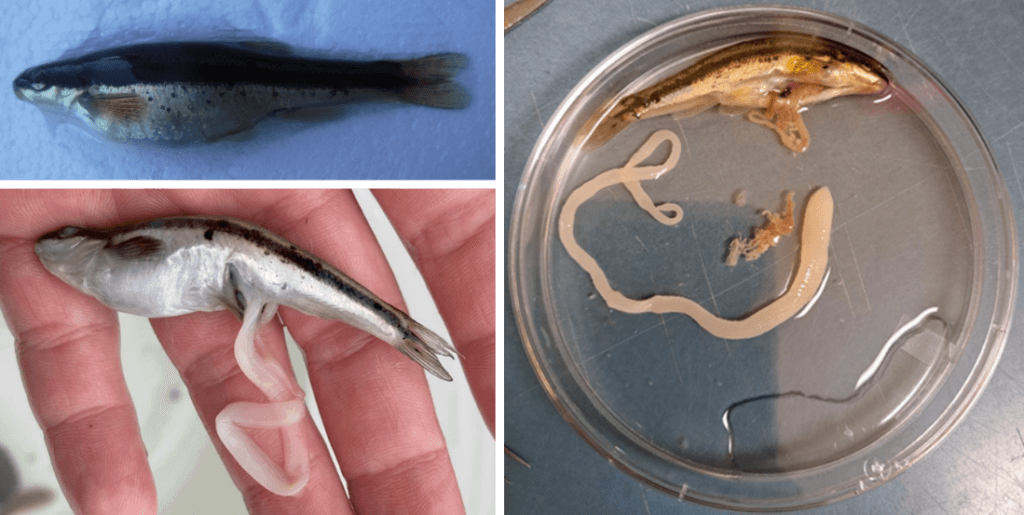
Notre utilisation des collections des pêcheurs récréatifs réduira la nécessité de sacrifier des poissons supplémentaires pour nos propres études. Au cours du printemps et de l’automne 2023, j’effectuerai une étude de plusieurs lacs du Nouveau-Brunswick pour répondre aux questions concernant l’aire de répartition du parasite, les poissons hôtes permissifs et les tendances saisonnières de la prévalence de l’infection. Les indicateurs morphologiques de l’infection, tels que le gonflement abdominal ou l’éruption de ténias à partir de l’évent (l’ouverture par laquelle les œufs, le sperme et les déchets quittent le poisson), nous aideront à sélectionner les poissons à échantillonner (figure 3). Pour diagnostiquer un poisson atteint de cette infection, les méthodes antérieures nécessitaient l’euthanasie et la dissection. Nous modifierons une procédure de diagnostique non fatale et peu invasive de Berger et Aubin-Horth (2018) pour nous permettre de diagnostiquer les poissons infectés sans les euthanasier. Notre objectif est de collecter des échantillons auprès des pêcheurs récréatifs et d’utiliser des techniques de diagnostique non fatales, afin de réaliser notre étude avec le moins de dérangement écologique possible. Nos études apporteront des informations précieuses au domaine de la parasitologie au Nouveau-Brunswick et aux travaux parasitologiques concernant Ligula intestinalis dans le monde entier !
Remerciements :
Je remercie le Dr Michelle Gray, Ben Andrews et Sarah Van De Reep pour avoir collecté les vers solitaires originaux en vue de leur identification. Sans leur intérêt et leur curiosité, je ne participerais pas à cet important projet de recherche. Je remercie le comité de supervision de ma maîtrise, le Dr Michael Duffy, le Dr Shawn MacLellan et le Dr Michelle Gray pour leurs conseils et leur encadrement. Je remercie également Kerstyn Dobbs, Hannah Lazaris-Decken, Nathanael Moore, Stephanie Scott et Tyler Lynn de l’UNBF pour leur aide en laboratoire, ainsi que Sarah Van De Reep, Becky Graham, Sara Plant, Allain Cassie et John Robinson à Parcs Canada pour leur aide sur le terrain.
Les ressources :
- Arme, C., & Owen, R. W. 1968. Occurrence and pathology of Ligula intestinalis infections in British fishes. The Journal of Parasitology, 54, 272–280.
- Berger, C. S., & Aubin-Horth, N. 2018. An eDNA-qPCR assay to detect the presence of the parasite Schistocephalus solidus inside its Threespine stickleback host. Journal of Experimental Biology, 221(9), jeb178137.
- Biswas, R., & Ash, A. 2021. Impact of Endocrine Disruption on Host-Parasite Interaction: A Digest from a Cosmopolitan Cestode Model Ligula intestinalis. In Proceedings of the Zoological Society. pp. 1-9. Springer India.
- Carter, V., Pierce, R., Dufour, S., Arme, C., & Hoole, D. 2005. The tapeworm Ligula intestinalis (Cestoda: Pseudophyllidea) inhibits LH expression and puberty in its teleost host, Rutilus rutilus. Reproduction, 130, 939-945.
- Fraser, M.L., M.A. Gray, K.D.R. Dobbs, B.I. Andrews, S. Van De Reep, M. S. Duffy. 2023. First reports of Ligula intestinalis and a Schistocephalus sp. infecting small-bodied fish in New Brunswick, Canada. Journal of Parasitology 109:288-295. DOI: 10.1645/22-127
- Hecker, M., & Karbe, L. 2005. Parasitism in fish—an endocrine modulator of ecological relevance?. Aquatic Toxicology, 72, 195-207.
- Jyrwa, D. B., Thapa, S., & Tandon, V. (2016). Helminth parasite spectrum of fishes in Meghalaya, Northeast India: a checklist. Journal of Parasitic Diseases, 40, 312-329.
- Loker, E., & Hofkin, B. 2015. Parasitology: a conceptual approach. Garland Science, Taylor & Francis Group, LLC. 711 Third Avenue, New York, NY, 10017, USA
- Loot, G., Aulagnier, S., Lek, S., Thomas, F., & Guégan, J. F. 2002. Experimental demonstration of a behavioural modification in a cyprinid fish, Rutilus rutilus (L.), induced by a parasite, Ligula intestinalis (L.). Canadian Journal of Zoology, 80, 738-744.
- Negrazis, L., Kidd, K. A., Erdozain, M., Emilson, E. J., Mitchell, C. P., & Gray, M. A. 2022. Effects of forest management on mercury bioaccumulation and biomagnification along the river continuum. Environmental Pollution, 310, 119810.
- Orr, T. S. C. 1966. Spawning behaviour of rudd, Scardinius erythrophthalmus infested with plerocercoids of Ligula intestinalis. Nature, 212, 736-736.
- Schabuss, M., Gemeiner, M., Gleib, A., Lewis, J. W., Miller, I., Möstl, E., & Grillitsch, B. 2005. Ligula intestinalis infection as a potential source of bias in the bioindication of endocrine disruption in the European chub Leuciscus cephalus. Journal of Helminthology, 79, 91-94.
- Szalai, A. J., Yang, X., & Dick, T. A. 1989. Changes in numbers and growth of Ligula intestinalis in the spottail shiner (Notropis hudsonius), and their roles in transmission. The Journal of Parasitology, 75, 571–576
- Trubiroha, A., Kroupova, H., Wuertz, S., Frank, S. N., Sures, B., & Kloas, W. 2010. Naturally-induced endocrine disruption by the parasite Ligula intestinalis (Cestoda) in roach (Rutilus rutilus). General and Comparative Endocrinology, 166, 234-240.
- Trubiroha, A., Wuertz, S., Frank, S. N., Sures, B., & Kloas, W. 2009. Expression of gonadotropin subunits in roach (Rutilus rutilus, Cyprinidae) infected with plerocercoids of the tapeworm Ligula intestinalis (Cestoda). International Journal for Parasitology, 39, 1465-1473.
- White, C. F. H. 2020. Fish effects from forest harvest and ectoparasitic copepods in northern New Brunswick. MScF Thesis, University of New Brunswick, 120 pp.
- Yoneva, A., Scholz, T., M?ocicki, D., & Kuchta, R. 2015. Ultrastructural study of vitellogenesis of Ligula intestinalis (Diphyllobothriidea) reveals the presence of cytoplasmic-like cell death in cestodes. Frontiers in zoology, 12, 1-9.



